Neurophysiologische Grundlagen des
Bewußtseins
1 Evolution und Aufbau des Gehirns
Entwicklungsgeschichtlich gesehen haben
sich von Insekten über Säugetiere, Affenvorläufer und Anthropoiden bis zum Menschen
alle Hirnregionen im Verhältnis zum Körpergewicht vergrößert. Vor allem der Anteil
des Neocortex (äußere Schichten) am Volumen des Gehirns hat in Richtung Homo Sapiens
zugenommen. Die relative Größe einer bestimmten Hirnregion ist dabei das Resultat
eines evolutionären Anpassungsprozesses der Spezies an eine bestimmte ökologische
Niesche. Die Größenzunahme der neocortikalen Rinde beim Menschen dürfte das Resultat
der abnehemenden Spezialisierung der Sensorik auf bestimmte Sinnesmodalitäten
und wenige Reaktionsmuster sein, die eine Ausprägung neuer Funktionen wie Sprache
(bzw. exakte zeitliche Steuerung und Speicherung von Tönen) sowie geistiger und
ästhetischer Fähigkeiten erlaubte. So hat sich der Neocortex vermutlich als Basis
für eine multisensorische Repräsentation der Umwelt entwickelt. 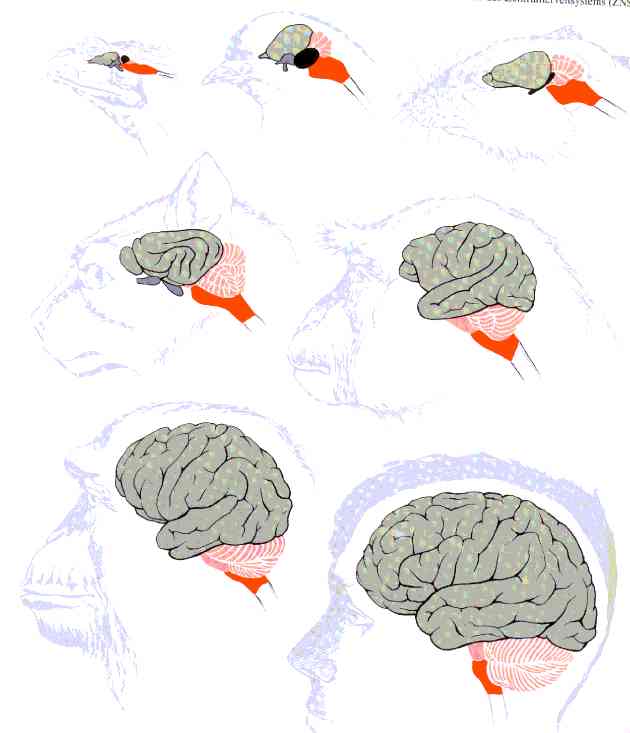
Fig1: Evolution cerebraler Strukturen
Das menschliche Gehirn wird in 5
Hauptabschnitten beschrieben:
* Neocortex, Basalganglien, Limbisches System
(zusammen das Endhirn. in Fig1 der obere und größte Teil des Gehirns)
* Thalamus, Hypothalamus (zusammen Zwischenhirn, zentral gelegen)
* Tectum, Tegmentum (Mittelhirn, zentral gelegen, unter dem Thalamus)
* Cerebellum Pons (Hinterhirn, in Fig1 gestreift dergestellt)
* Medulla oblongata (Nachhirn, in Fig1 nahe dem Wirbelkanal)
Diese Hauptabschnitte werden im Embryo
bereits 6 Wochen nach der Befruchtung sichtbar. Funktionell nimmt die Flexibilität
und Geschwindigkeit der Informationsverarbeitung nach außen hin zu (Richtung Neocortex)
Bewußtseinstechnisch interessant sind von den unteren Hirnschichten vor allem
der Hypothalamus und der Thalamus. Der Thalamus fungiert als Relaissystem für
die Weiterleitung von sensorischen Informationen zur Hirnrinde und ist daher das
Tor zur Aufmerksamkeit. Außerdem dient er als "Takt- und Impulsgeber" für die
rhythmischen elektrischen Aktivitäten des Großhirns. Der Hypothalamus ist in enger
Verbindung mit dem limbischen System an der Steuerung von Lernvorgängen beteiligt.
Das limbische System selbst beinhaltet das Geruchszentrum (Bulbus olfactorius),
das Funktionen für Lernen (Einprägen) und kognitive Prozesse unterstützt. (besonders
deutlich bei Geruchs- und Geschmacksaversionen). Das limbische System wird so
zur Schaltstelle für informationsverarbeitende Prozesse und Kurzzeitgedächtnis
und bildet die anatomischen Grundlagen für Emotionen. Es ist neben dem Hypothalamus
stark mit dem frontalen Assoziationscortex verbunden.
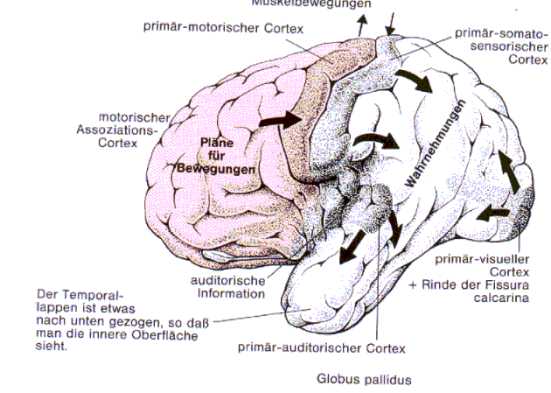
Fig2: Überblicksmäßige Darstellung der primären sensorischen und motorischen Areale
des Neocortex, Birbaumer 1991
Bei Fig3 wird eine elektrische Reizung
des Gehirns von außen gezeigt. Interessanter Effekt des Anlegens eines externen
Spannungspotentials (5V, 12mA) ist, daß auf der Seite der anodalen (+) Polarisierung
eine Anregung des Verhaltens entsteht. (Begünstigung von Entscheidungen, die diese
Hemisphäre betreffen, Erleichterung von informationeller Verarbeitung)

Fig3: Verteilung der Sensomotorischen Regionen über den vertikalen
Querschnitt der Hirnrinde, Birbaumer 1991
2 Analyse und Messung von Gehirnaktivitäten
Einige Methoden, um auf Funktion oder
Aktivität im Gehirn zu schließen:
- EEG, MEG (Magnetoencephalogramm)
- Färbung und Elektronenmikroskopie
- Stereotaxie: Einstechen einer Elektrode an den zu untersuchenden Punkt im Gehirn
- Läsion: Bewußte Zerstörung von Hirnregionen (bei Menschen temporär)
- Reizung durch Hochfrequenz oder elektrische Ströme (siehe Fig3)
- Röntgencomputertomographie (Cat-Scan)
- Positron-Emissions-Tomographie bzw Messung regionaler Hirndurchblutung
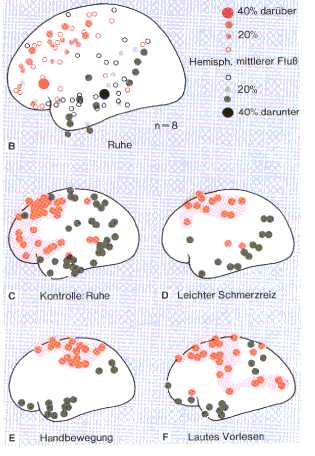
Fig4: Messung der regionalen Hirndurchblutung
mittels intraarterieller Injektion von Xe133 in die Halsschlagader, Birbaumer,
1991
2.1 Elektroencephalographie und Ableitung
von Gehirnwellen
EEG ist die einfachste und humanste
Methodik, um Gerhirnaktivitäten zu analysieren, aber nicht immer ausreichend genau,
da die Abnahme der elektrischen Potentialsveränderungen der Großhirnrinde und
der unterliegenden subcortikalen Potentiale an der Oberfläche erfolgt und so auch
die umliegenden Bereiche in die Messung eingehen, bzw. Artefakte (durch Augen-
oder Muskelbewegungen) die Messung stark beeinflußen. Daten einer großen Anzahl
von Elektroden geben einen guten Überblick und können im Computer zu einem 3-dimensionalen
Bild der Hirnaktivitäten zusammengefügt werden. Spannungsschwankungen, die von
außen gemessen werden, bewegen sich in Bereichen von 1 Mikrovolt (hohe Betafrequenzen)
bis zu 300 Mikrovolt (Epileptischer Anfall). Man unterscheidet unipolare (1 Elekrode
gegen eine inaktive Region wie Kinn oder Ohrläppchen) von bipolarer Ableitung
(Spannungsdifferenz 2er aktiver Regionen)
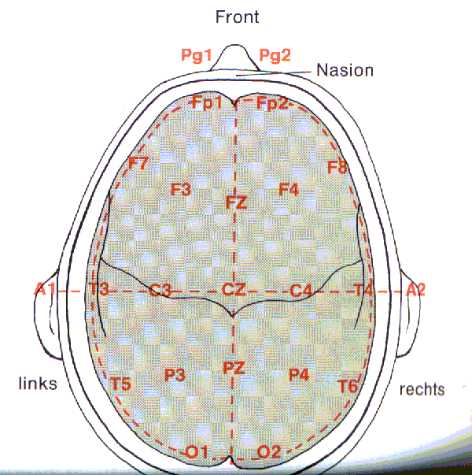
Fig5: internationales 10-20-System
zur Ableitung des EEG, Birbaumer 1991
2.2 Analyse der Daten
Vor allem durch computerunterstützte
Verarbeitung ist eine Analyse des EEGs einfach und schnell (real-time) möglich.
Man erhält Aufschlüsse über Aktivitäten in den verschiedenen Frequenzbändern,
Koherenz-Analysen und Gleichspannungsschwankungen (siehe 2.2.2 EKP bzw 2.2.3 LP)
2.2.1 Frequenzbänder
Die Frequenz beim EEG reicht von Gleichspannungsschwankung
(0Hz) bis über 100 Hz. Bewußtseinstechnisch interessant sind vor allem die unteren
Bereich der Frequenzen, die unterteilt sind in Delta, Theta, Alpha und Beta. Synchronisationseffekte
in niedrigen Frequenzbereichen haben Entsprechungen im Verhaltenskontinuum bzw.
Bewußtsein:
DELTA (unter 4 Hz) : Schlaf (tiefer Slow-Wave-Sleep)
THETA (4 - 8 Hz) : Visualisieren, Kindheitserinnerung bzw. Wach-Träume, dösen, verminderte
Reaktion auf die Außenwelt
ALPHA (8 - 13 Hz) : entspannter Wachzustand, kreative Gedanken, gute Kommunikationsfähigkeit
Bewußtseinstools wie MindMachines nützen diese Effekte und versuchen das Gehirn
mit wählbaren Frequnzbereichen zu synchronisieren, um zB einen Theta-Zustand,
der normalerweise nur kurz vor dem Einschlafen bewußt erlebt wird, länger aufrechtzuerhalten.
Sehr starke Aktivierung (Stress, emotionale
Erregung) zeigt sich in desynchonisierten, niedrigen Amplituden. Wache Aufmerksamkeit
hat eine Entsprechung in einem gemisch schneller Frequenzen.
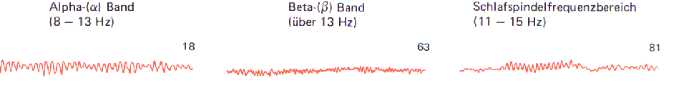
Fig6: Alpha- und Betaaktivität bzw
Schafspindeln, Birbaumer 1991
2.2.2 EKP
Unter Ereigniskorrelierten Hirnpotentialen
versteht man alle elektrocortikalen Potentiale, die vor, während oder nach einem
sensorischen, motorischen oder psychischen Ereignis im EEG meßbar sind. EKPs sind
in der Regel von sehr viel kleinerer Amplitude (1 - 30 Mikrovolt) als das Spontan-EEG,
das diese Potentiale als Rauschen stark überlagert. Der Grund für die kleinen
Amplituden der EKPs liegt sowohl in ihrer stärkeren örtlichen Lokalisation in
den verschiedenen Cortexarealen, die eine Konsequenz der lokalen thalamischen
Generatoren ist, als auch in der Tatsache, daß sie seltenere Ereignisse sind als
die in Form und Amplitude ähnlichen, dauernd vorhandenen EEG-Wellen. EKPs werden
durch die Mittelungstechnik aus dem überlagernden Rauschen herausgefiltert, indem
man davon ausgeht, daß bei der Wiederholung eines Reizes ähnliche EKPs produziert
werden, die dann summiert und durch die Anzahl der Messungen dividiert werden.
Es kristallisiert sich somit die Form des EKPs heraus (unter der Annahme, daß
die übrigen Signale des EEGs zufällig verteilt sind). Es wird nun versucht, aus
den EKPs auf unterschiedliche neurophysiologische und damit psychologische Vorgänge
zu schließen, die durch den Reiz hervorgerufen werden. So haben zB. Amplitudenveränderungen,
die bis zu 10ms nach dem Reiz hervortreten, ihren Ursprung im Hirnstamm und werden
von dort bis zur Schädeldecke fortgepflanzt. Komponenten zwischen 10 und 100ms
nach einem Reiz entstehen zum Großteil in den spezifischen Projektionsarealen
des Neocortex, ihre Bedeutung für die Informationsverarbeitung ist aber nicht
völlig klar. Ab 100ms zeigen die Komponenten Variationen von psychischen Veränderungen,
die als Reaktion auf den Reiz auftauchen und sind nicht mehr von den Reaktionsbedinungen
alleine abhängig, sie werden daher als endogene Komponenten bezeichnet.
2.2.3 LP
Langsame Potentiale sind Gleichspannungsverschiebungen
des EEG in negative oder positive Richtung, die eher träge reagieren (200 - 300
ms), und die Aktivitäten eines ausgedehnten neuronalen Systems spiegeln. LPs spielen
in der Psychologie eine große Rolle, da sie für die Planung und Mobilisierung
zielgerichteten Verhaltens notwendig sind. LPs im negativen Bereich treten stets
dann auf, wenn zusätzliche Energiereserven in neuen komplexen Situationen oder
psychischen Bedingungen benötigt werden, LPs im positiven Bereich durch die Hemmung
oder Konsumation der Mobilisierung. In einer Versuchsanordung wird einer Person
ein Signalton präsentiert, auf den 6 sek. später ein weiterer akustischer Reiz
folgt, auf den die Person so schnell wie möglich eine Taste drücken muß. Die entstehenden
LPs wurden mit Komponetenanalysen verschiedenen Hirnregionen und Verarbeitungsprozessen
zugeordnet. Die erste, frühe negative Komponente kommt aus dem präfrontalen Cortex,
während die späte Komponente stets in jenen Hirnregionen dominiert, in denen der
zweite Reiz verarbeitet oder die Reaktion vorbereitet wird. (zB. bei Reaktion
mit der linken Hand ein lokales Maximum über der rechten pracentralen Windung)
Bis 300ms sind einzelne Verarbeitungsschritte
nicht bewußt, bewußtes Erleben ist im allgemeinen an hinreichend synchrone Aktivierung
eines größeren Zellareals gebunden. In der Regel treten klar feststellbare Bewußtseinsänderungen
erst nach der ersten LP-Komponente (auch CNV genannt, contingente negative Variation).
Während die Negativierung bis 100ms im wesentlichen auf das primäre Projektionsareal
beschränkt bleibt, breiten sich die übrigen Potentialanteile in verschiedenen
Hirnregionen aus, je nach den oft weit auseinanderliegenden Arealen, die an einem
bestimmten Verarbeitungsschritt beteiligt sind.
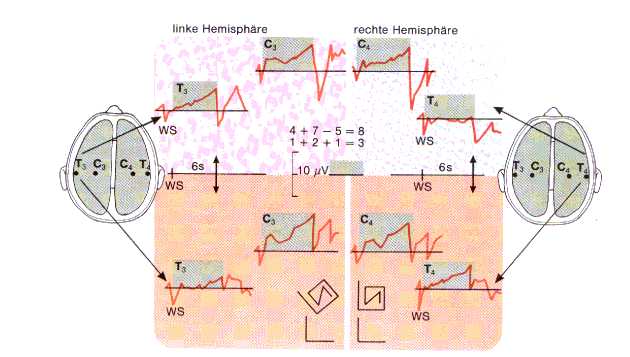
Fig 8: LPs bei Aufgaben, Birbaumer
1991
In Fig8 oben arithmetische Aufgaben,
unten Erkennen von verdrehten Figuren. Die Aufgaben wurden nach einem 6 sek. dauerden
Vorintervall dargeboten. (WS). Vor der Darbietung der Aufgaben bildet sich eine
antizipatorische Negativierung aus, bei Darbietung der Aufgaben kommt es zu einer
Positivierung. Signifikante Unterschiede treten nur in den temporalen Ableitungen
auf: Bei Rechenaufgaben negativiert die linke Hemisphäre stärker als die rechte
(T3 und T4 oben), bei Gestaltungsaufgaben negativiert die rechte Hemisphäre stärker
als die linke (T3 und T4 unten)
Durch Biofeedback können Versuchspersonen
die Aktivierung von LPs in verschiedenen Hirnregionen lernen. Wenn dann die Person
eine negative Potentialsveränderung über der erlernten Position erzeugt, reagiert
sie effizienter auf Aufgaben, die in diesem Areal verarbeitet werden.
3. Bewußtsein und Aufmerksamkeit
Die Produktion von Bewußtsein ist eine
Eigenschaft des Kurzzeitgedächtnisses (KZG, engl. STM), während Prozesse im LZG
in der Regel nicht bewußt sind. LZG-Inhalte werden erst bei der Übertragung ins
KZG bewußt. Jeder Gedächtnisinhalt im KZG erhält eine damit assoziirte Anreizkomponente,
die die Selektion und Speicherung des Inhaltes beeinflußt. Bewußtseinsprozesse
resultieren stets aus vorbewußter Informationsverarbeitung. Die Annahme eines
einzigen Selektionssystems ("Flaschenhalstheorie") ist mit den experimelntellen
Befunden nicht vereinbar. Alle Theorien der Aufmerksamkeit gehen von einer limitierten
Aufmerksamkeitskapazität aus. Der gemeinsame Mechanismus hinter allen Bewußtseinsformen
und Aufmerksamkeit wird in jenen Situationen sichtbar und meßbar, bei denen die
Anforderungen die Kapazität der Person für die Aufgabe überschreiten. Nur ein
Bruchteil der ankommenden Reize wird bewußt. Bewußtsein tritt nur auf :
- beim Erwerb neuer Information
- bei Abgabe von Urteilen und Wahlreationen
- bei Nicht-Eintreffen erwarteter Reize
3.1 Vorbewußte Informationsverarbeitung
Die Reizaufnahme, die Repräsentation
(Codierung) des Reizes, die Musterextraktion und der Vergleich des gegenwärtigen
Reizmusters mit Reizmustern im LZG sind ebensowenig bewußt wie die Auswahl und
Ausführung der auf den Reiz passenden Reaktion und die Rückmeldung des Reaktionserfolges
aus der Peripherie. Die Ressource eines KZG-Systems ist limitert auf 2-2,5 Bit
kann aber durch chunking, d.h. geeignete Kombination in Gruppen im Laufe eines
Lernprozesses erweitert werden.
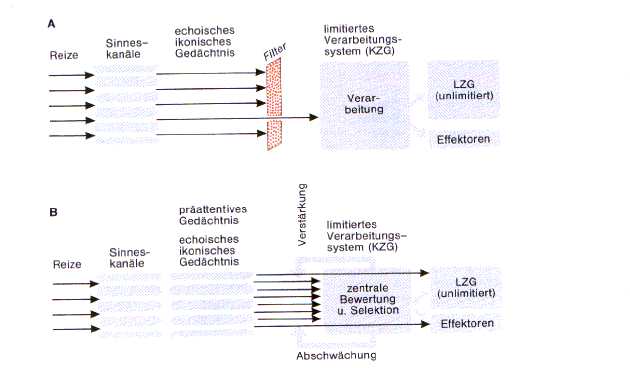
Fig9: Flaschenhalsmodell der Aufmerksamkeit
und Modell einer zentralen Informationsverarbeitung, Birbaumer, 1991
Flaschenhalstheorien besagen, daß aus
der Vielzahl der vorhandenen Reize der stärkste ausgewählt wird (zB der lauteste
Redner), der dann den selektiven Filter passiert und weiter verarbeitet wird.
Daß dies eine unvollständige Annahme ist, zeigt sich zB. durch Wahrnehmung des
eigenen Namens in einer lauten Gesellschaft etc.
Die Intensität einer Aufmerksamkeitszuordnung
hängt von einem Vergleichsprozeß zwischen ankommenden Reizmustern und im LZG gespeicherten
Modellen desselben Verarbeitungssystems (zB visuell, akustisch etc.) ab, die automatisert
erfolgt und in Fig. 7b durch die Pfeile, die direkt ins LZG führen, symbolisiert
wird, ab. Jede Abweichung vom gespeicherten Modell löst eine Orientierungsreaktion
(OR) aus. (200-250ms nach dem Reiz, Schwelle von unbewußter zu bewußter Informationsverarbeitung)
Orientierungen sind unauflöslich mit der Bildung von Erwartungen verbunden, die
das Resultat von zunehmenden Präzisierungen des im LZG gespeicherten Reiz-Reaktions-Modells
sind. Beim Auftreten eines bekannten Reizes wird dieser Inhalt vom LZG auf das
KZG projiziert. Paßt der im KZG aus den Sinneskanälen stammende Inhalt auf die
aus dem LGZ projizierten Inhalte, so wird die Erwartung bestätigt, sonst wird
eine Orientierung eingeleitet bzw. der Inhalt des LZGs modifiziert. Als Habituation
bezeichnet man die Verringerung der Intensität der OR nach wiederholter Darbietung
des Reizes.
3.2 LCCS-limited capacity control
system
ist ein hypothetisch angenommenes System
zentraler Aufmerksamkeitsmechanismen. Bei überlernten, geübten Aufgaben (zB autofahren,
wenn genug Praxis besteht) erfolgt die Reaktion ohne Bewußtsein, andere Reaktionssysteme
können gleichzeitig ohne gegenseitige Behinderung funktionieren. Die gesamte ankommende
Information wird zunächst für wenige ms in einem sensorischen Speicher gehalten.
dort wird Mustererkennung, Codierung und danach ein Vergleich (match) vorgenommen.
Paßt der angekommene Reiz vollständig in ein überlerntes Reiz-Reaktions-Muster,
wird die Reaktion "automatisch", d.h. ohne besondere Erhöhung der Erregung, in
den beteiligten Netzwerken und ohne Bewußtsein ausgelöst.
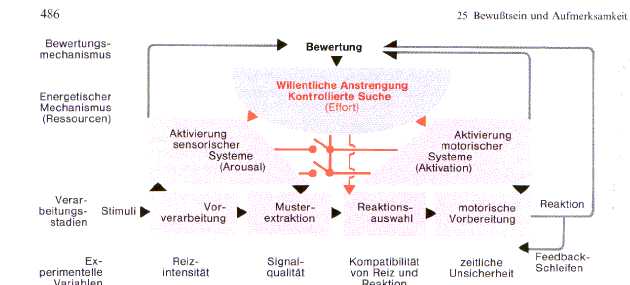
Fig10: Kognitive Prozesse der Informationsverarbeitung,
Birbaumer 1991
Erst wenn neue und komplexe Situationen
und Handlungen auftauchen, erregt das LCCS zusätzlich informationsverarbeitende
und reaktionsplanende Systeme und hemmt die nicht-beteiligten. Reize und Reaktionen,
die in der Vergangenheit mit biologisch bedeutsamen Reizen (zB. Triebbefriedigung)
assoziirt waren, lösen eher einen Erregungsanstieg aus. (Effort-Mechanismus, geht
mit Bewußtsein und verstärktem Energieverbrauch (zB Glucose) einher)
Wenn Überlappungen zwischen den auf
Fig11 dargestellten Verarbeitungsdimensionen durch simultane Aufgabendarbietung
entstehen, wird das LCCS aufgerufen und die Aufmerksamkeitsenergie auf eine Dimension
(zB visuell) verteilt (Ressourcen-Zuordnung). Jedes der gezeigten Verarbeitungssysteme
verfügt über beschränkte Ressourcen, sonst könnten mehrere Aufgaben innerhalb
einer oder auch in verschiedenen Verarbeitungsdimensionen ohne Interferenzen parallel
gelöst werden. Dies gelingt innerhalb einer Dimension am schlechtesten (neue akustische
Aufgaben stören akustische mehr als visuelle)
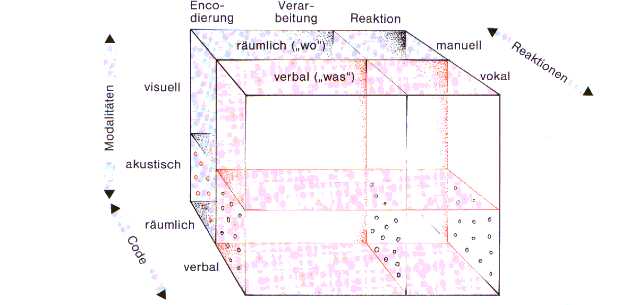
Fig11: einige wichtige Verabeitungsmechanismen,
Birbaumer 1991